
どうやったら物質から生命が生まれるのか?
研究の背景
未解決問題
近年の科学の発展は目覚ましいものがあります。寿命はますます長くなり、巨大なエネルギーや情報を扱えるようになり、人類はどんどん丈夫で強く賢い生物になっていっているように思えます。
しかし、私たちは未だ生命がどうやって生まれたのかを理解していません。現在の生物学では生物は生物からしか生まれないことになっていますが、少なくとも一度は無生物から生物が生まれなければ生物は存在できません。いったいどうやって物質から生命が誕生しうるのか? これが人類に残された大きな未解決問題の一つです。
この問題に対する答えは「進化」だと予想されています(1)。つまり、原始地球のどこかの段階で自己複製して進化する能力を持った単純な分子(あるいはその集合体)が生まれ、それが進化することによって現在見られるような複雑な生命になって行ったと予想されています。
しかし、この予想は完全に想像の産物です。いままで誰も単純な分子が進化して複雑な生命体へと近づいて行った様子を見たことはありません。むしろ後述するように、試験管の中で進化するシステムを作ってみても、より単純化する方向へ進化していきます(2)。
それではどうやったら単純な物質から複雑な生命が生まれるのでしょうか? おそらく、ただ進化するだけでは不十分で、何か別の条件が整わないと生命を生み出すような進化は起きないのだと私たちは考えています。その条件を未だ私たちは理解していないのでしょう。それゆえに私たち人類は未だに自らの出自をも理解していない根無し草のような状態にあります。私たちはどこから来たのかもわからず、おそらくそれゆえにどこへ向かっているのかもわかっていないように思えます。
私たちの戦略
私たちの研究室では、人類に残された未解決問題である「どうやって物質から生命が誕生しうるのか?」を(少なくとも一部でも)解き明かしたいと考えています。そして、物質から生命を生み出した進化という現象の特性を理解したいと考えています。そのための私たちの戦略は「進化実験」です。どうやって生命が誕生するのかを理解するために最も効果的な方法は、実際に分子を組み合わせたものを試験管の中で進化させてやって、生命らしさを獲得していく様子を直に観察することです。試験管の中でならば、進化途中の様子もすべて調べることができます。途中から環境を変えてやり直すこともできます。生命誕生に必要な条件をあいまいさなく理解できるはずです。
過去に初めて試験管内で分子の複製と進化を行ったのはアメリカのシュピーゲルマンらのグループです。彼らは1967年にウイルスゲノムに由来するRNAと、同じウイルスのRNA複製酵素(こちらは精製したものを入れています)を使って、RNAの持続的な複製と進化を初めて試験管内で起こすことに成功しました(2)(図1)。しかし、この実験では確かに突然変異による多様性の出現と自然選択によるダーウィン進化はおきたものの、その進化は複雑な生命を生み出すものとは逆で、RNAはどんどん短く単純な方向へ進化していってしまいました。最終的に集団を占めたRNAは複製酵素によって複製される能力だけを持つ500塩基ほどの短いRNAでした(シュピーゲルマンモンスターと呼ばれたりもします)。後の実験でさらに進化を続けた人もいますが、RNAはもっと短くなっていくだけで(90塩基くらいにまで短くなります)、生命体が出現する気配すらありません。その後も複数のグループがRNAやDNAと酵素を組み合わせて自己複製して進化する分子システムを構築していますが、単純化方向への進化が起こり、生命に近づいていく様子が観察されることはありませんでした(3)。そのために分子を使った進化実験は、90年代以降はほとんど進展はなく、酵素や核酸の改良という応用方面への利用にとどまっていました。
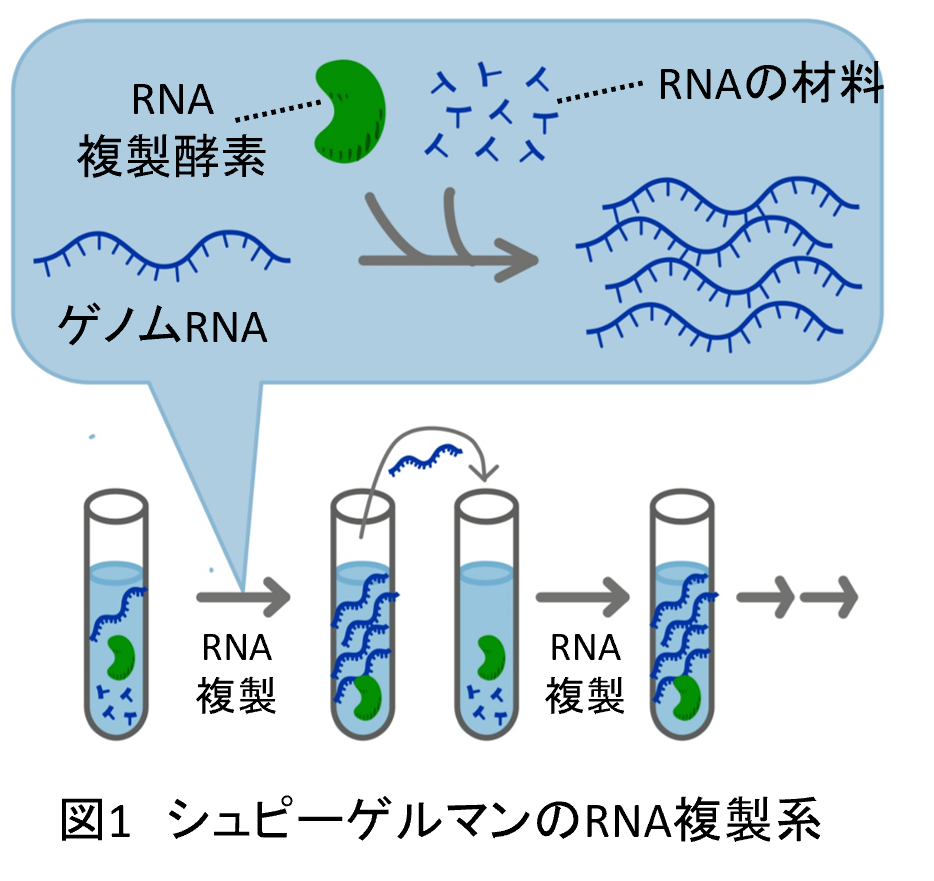
なぜ、これまで作られていた分子の自己複製システムは進化させても生命に近づいていく様子はなく、むしろ単純化してしまうのでしょうか? 何が足りないのでしょうか? 一つの可能性は、遺伝情報分子と機能分子の分離です。現在の生物では遺伝情報はDNAという分子に(一部のウイルスの場合はRNA)、機能はタンパク質という分子に分けています。この分離が生命誕生の条件の一つだと理論的な考察がされています(4)。
そこで私たちはこの情報分子と機能分子の分離を、シュピーゲルマンらのRNA複製システムに組み込みました。つまり、シュピーゲルマンらのシステムでは研究者がそとから入れてやっていたRNA複製酵素を、情報(遺伝子)としてRNA自身に載せておき、RNAから複製酵素が翻訳されて、その複製酵素によってRNAが複製されるようにしました(図2)。このシステムの構築には、RNAから複製酵素タンパク質を翻訳させる仕組みが必要になりますが、折しもそのころ東大の上田卓也研究室から既知の分子の組み合わせで翻訳を達成するシステムが開発されました(5)。このシステム(PURE systemと呼ばれます)を使うことでRNAが自分の情報を使いながら複製し進化するシステムを作ることができました(6)(注1, 注2)。下に示すように、このシステムであればこれまでの分子進化システムでは起きず、そしてこれまで生物の進化でしか起きなったような様々な進化現象が起きるようになりました。私たちの研究室では、このRNA複製システムを実験モデルとして用いて、生命らしさが進化する条件の探索と、進化という現象の理解を目指しています。
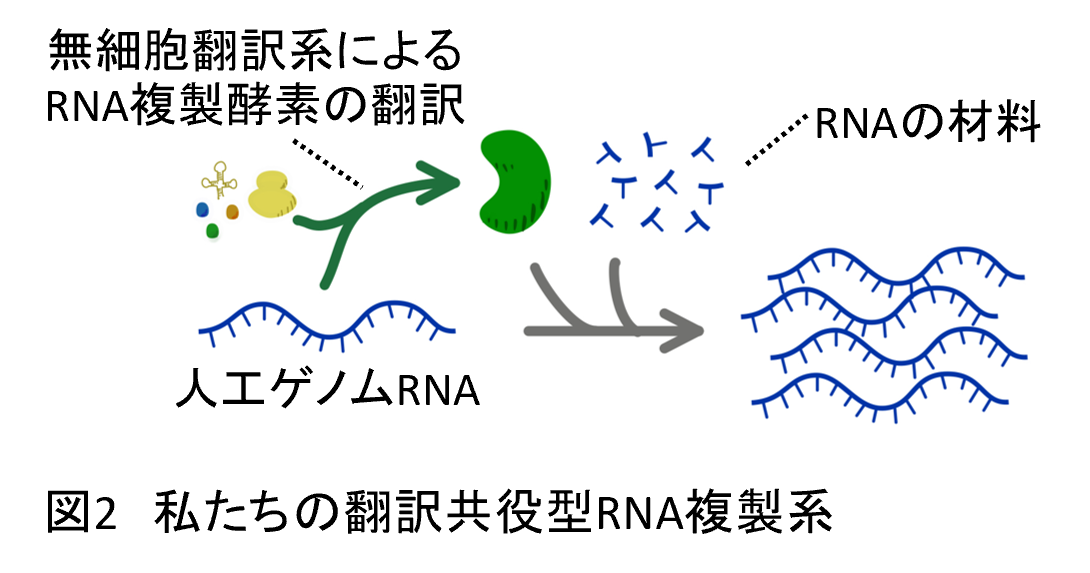
注1: いったいどうやって翻訳機構が生まれたのか?という問題は大きな未解決問題として残っています。また、PURE systemは原始地球に存在したはずはないのに、そんなものを使っていいのかと思われる方がいるかと思います。こういった批判に対して市橋は「どうやって物質から生命が誕生しうるのか?」という問題と「原始地球でどうやって翻訳機能が生まれたのか?」という問題は分けて答えるべきだと考えています。
生命の起源は極めて大きな問題です。最初の生命が何からできていたのか、その材料はどうやってできたのか、どこで生まれたのか、どうやって今の生物にまで進化できたのか、何から何までわかりません。しかも、地球科学、化学、生物学など複数の分野にまたがった問題です。このような問題を解くためには質問を分ける必要があると考えます。生命の起源の研究者の多くは原始地球における有機物質の起源を研究されていますが、そのような原始地球における有機物質の起源の問題と、有機物質が生まれた後の自己複製体の起源の問題は分けて解くことができるはずです。さらに自己複製体がうまれてからどうやって情報分子と機能分子の分離が起きたのかの問題も分けられます。さらに情報分子と機能分子の分離が起きた原始生命体がどうやって現在のような生物へと進化したのかも別の問題として切り分けられます。私たちはこの最後の問題に取り組んでいます。それはこの問題が現在の私たちの実験モデルで解明できる問題であるのと同時に、この問題こそが物質から生命への遷移を可能とした最も重要な問題だと信じるからです。そして、この問題を解くためには、特に原始の材料を使う必要はありません(そもそもそれがどんなものだったのはわかりませんし)。ただ生きていないものの組み合わせであれば、そこからどうやって生命らしさが生まれるのかを理解するには十分です。
注2: 翻訳システムの導入により、もう一つ細胞のような区画構造が必要になりました。これがないとランダム変異の蓄積により、複製が持続しなくなります。なぜ必要になるかはこちらを参照ください。私たちの実験系では、これを油中水滴(油の中に分散させた微小水滴)で実現しています。現在、地球上で見つかっているすべての生物は細胞から構成されていますが、そのような構造は、すでにこのレベルの複雑さを持つ自己複製分子の時代から必要だったと考えられます。
参考文献(*は当研究室からの論文)
-
Joyce, G. F. The Antiquity of RNA-Based Evolution. Nature 2002, 418, 214-221
-
Mills, D. R.; Peterson, R. L.; Spiegelman, S. An Extracellular Darwinian Experiment with a Self-Duplicating Nucleic Acid Molecule. Proc. Natl. Acad. Sci. USA 1967, 58, 217-224
-
Wright, M. C.; Joyce, G. F. Continuous in Vitro Evolution of Catalytic Function. Science. 1997, 276, 614-617; Breaker, R. R.; Joyce, G. F. Emergence of a Replicating Species from an in Vitro RNA Evolution Reaction. Proc. Natl. Acad. Sci. 1994, 91, 6093-6097
- Ruiz-Mirazo, K.; Umerez, J.; Moreno, A. Enabling Conditions for “Open-Ended Evolution.” Biol. Philos. 2008, 23, 67-85
-
Shimizu, Y.; Inoue, A.; Tomari, Y.; Suzuki, T.; Yokogawa, T.; Nishikawa, K.; Ueda, T. Cell-Free Translation Reconstituted with Purified Components. Nat. Biotechnol. 2001, 19, 751-755
-
*Ichihashi, N.; Usui, K.; Kazuta, Y.; Sunami, T.; Matsuura, T.; Yomo, T. Darwinian Evolution in a Translation-Coupled RNA Replication System within a Cell-like Compartment. Nat. Commun. 2013, 4, 2494.
これまでの成果
寄生体がいない条件での進化実験
まず私たちは翻訳システムを導入したRNA複製システム(以後RNAの自己複製システムと称します)を微小水滴の中で長期間継代してみました(1)。継代の方法は、まず最初のRNAを翻訳反応液(PURE system)に入れ微小水滴化してからて37℃で2-5時間温めます(図3)。そうすると水滴内でRNAから複製酵素の翻訳とそれによるRNAの複製が起こりますが、そのうち基質(多くの場合NTP)を使い切って翻訳と複製が止まります。その後、一部の水滴をとって新しいPURE systemを含む微小水滴と混ぜます。このとき微小水滴どうしの融合と分裂が起こり、RNAに新しい基質と翻訳に必要な因子が供給されます。この水滴を温めると、再び複製酵素の翻訳とそれによるRNA複製が起こります。こうしてPURE systemを含んだ水滴を供給しながら継代することでRNAの翻訳と複製を永遠に続けることができます。
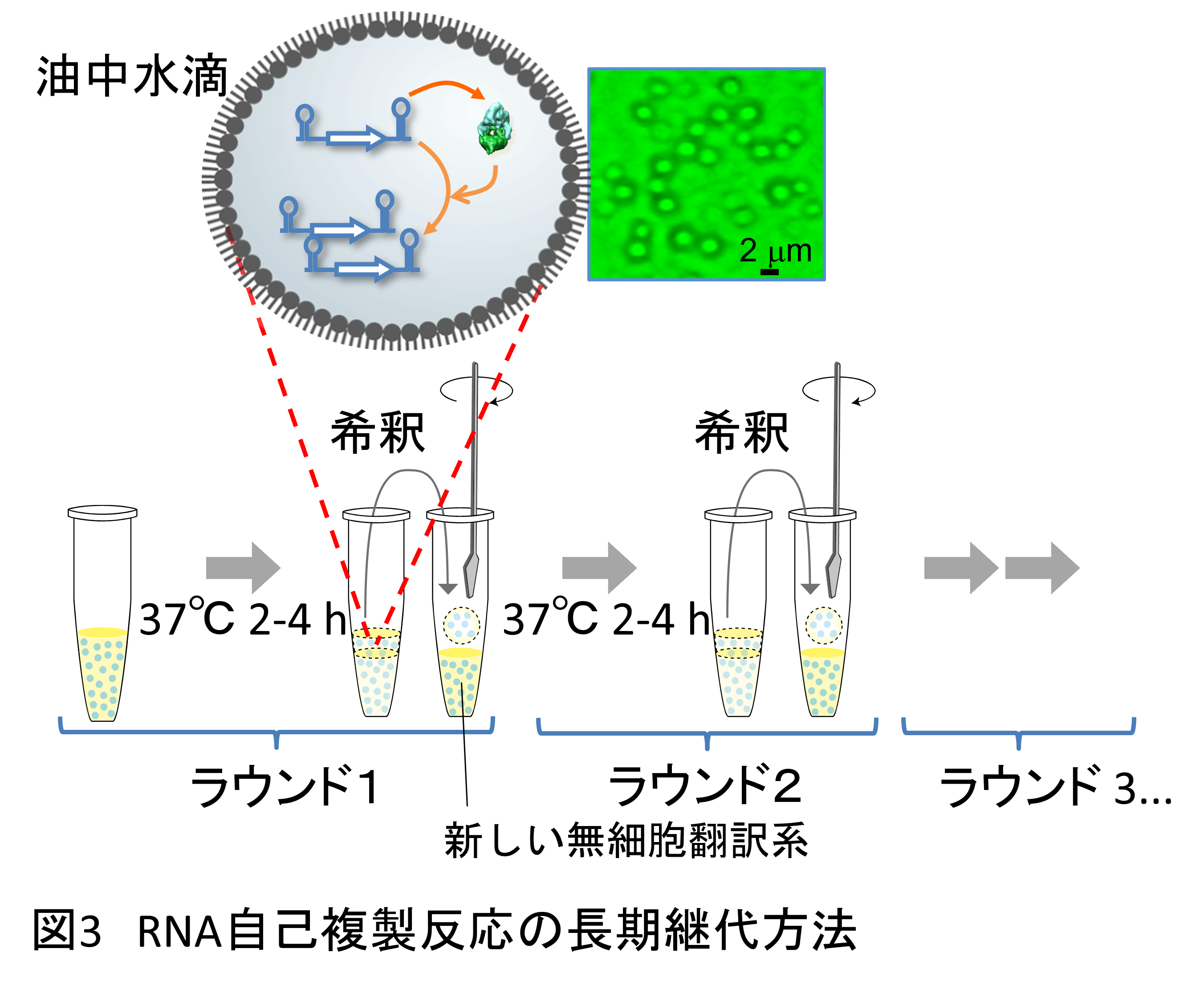
この継代をずっと繰り返していくと、そのうちRNAに突然変異が入ります。これは主にRNA複製が完璧ではなく時々間違えてしまうことによります。そしてもし、変異の入ったRNAがもとのRNAよりも複製能力が高くなると、その変異型RNAは集団内で割合を増やしていくことになります。こうしてダーウィン進化(適応進化)が起こるはずです。
私たちはこの継代反応を約120ラウンド行いました。その結果、RNAに変異が徐々に蓄積し、それに伴いRNAの複製能力が向上していくことを見出しました。(図4)。つまり複製能力の高いRNAが進化しました。シュピーゲルマンらの実験とは異なり、この進化ではRNAはほとんど短くなっていませんでした。それはもし短くなると複製酵素遺伝子が削られてしまい、単独では複製できなくなってしまうから当然です。したがって、この実験で初めて、単純化せずに分子システムを進化させることに成功したということになります。
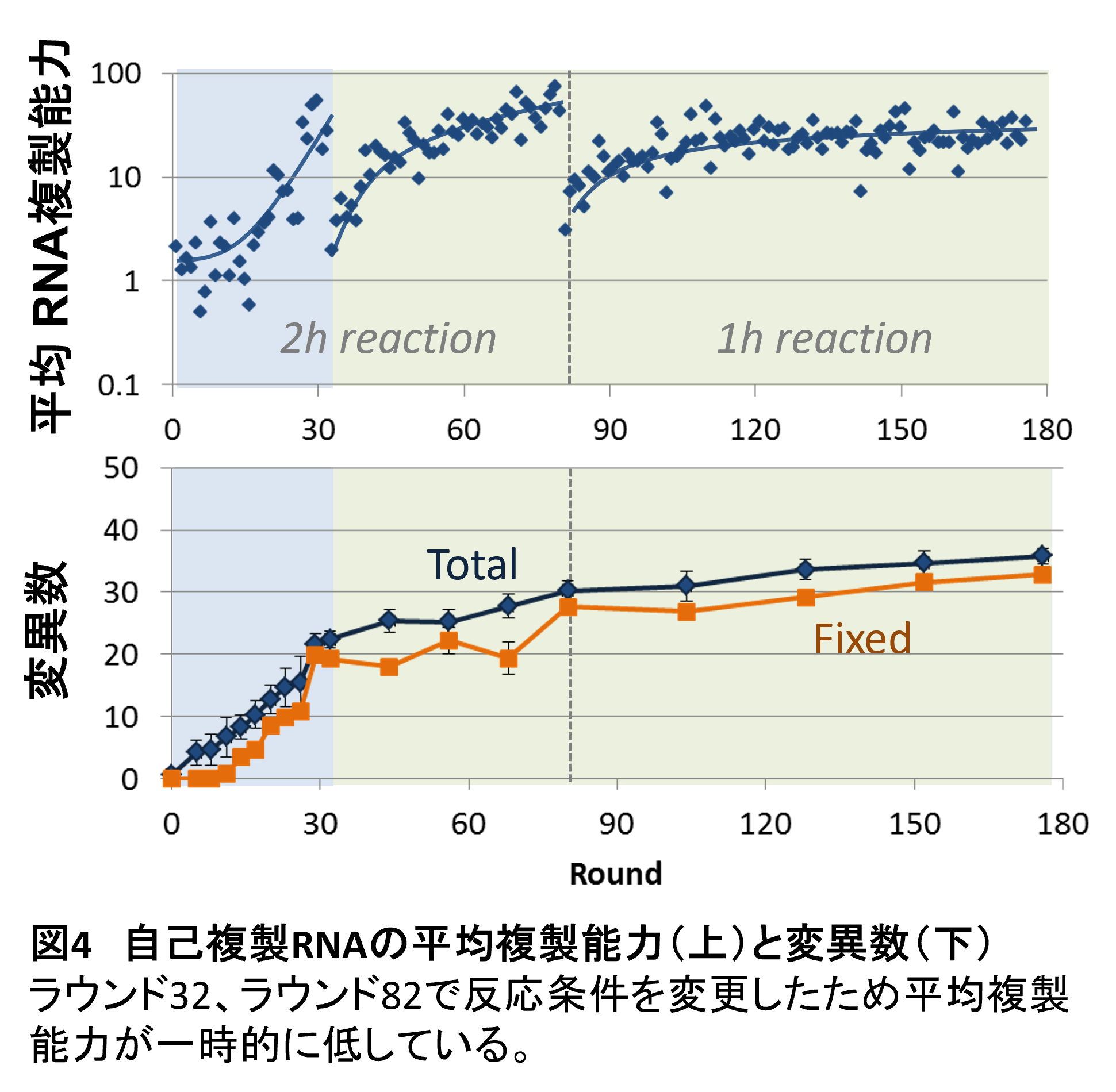
ただ、ではこのままこのRNAの進化を続けていくと、生命へと近づいていくのかと言えば、とてもそうは思われませんでした。まず進化はだんだん遅くなっていっています。100ラウンドの継代を超えると、もうほとんど変異も集団内に固定されず、複製能力もほとんど上がらなくなりました。つまり進化はほとんど止まってしまったように見えました。この「進化がすぐ止まる」というのも自己複製分子の進化の特徴の一つです。過去の進化実験においても進化は思ったより早く起きますが、すぐに最適値にたどり着いて止まってしまいます。おそらく適応度地形が極めて多峰性で局所的な最適値がそこら中にあるのではないかと考えています。
さらに生物進化の特徴である多様性も現れません。この進化実験の系統樹を書いてみると、変異率が高いために細かい枝分かれはありますが、基本的に1本の幹を維持したまま伸びていきます(図5)。後に図10で示すような多様なRNA種に分化するようなことは起きませんでした。

寄生体がいる条件での進化実験
そこで次に私たちはRNA自己複製システムに寄生体となるようなRNAを導入することにしました。実は、そのようなRNAは継代中に自然発生していました。このRNAは複製酵素遺伝子を持たず、複製酵素によって認識される配列だけは維持しているような短いRNAです(図6)。この短いRNAは複製酵素遺伝子を持たないため単独では増えられませんが、同じ水滴内に普通のRNAがいれば、そこから翻訳された複製酵素を奪って増えることができます。この意味でこの短いRNAはもとの長い(複製酵素遺伝子を持った)RNAを宿主としてそこに寄生するRNAです。このようなRNAは複製酵素遺伝子をもったRNAからRNA組換えによって一定確率で生まれてきます(注3)。
RNAは低い頻度(10の-7乗/hくらい)で自発的に組換えを起こすことが知られています。普通の実験では検出できない頻度ですが、私たちの反応系ではRNAの分子数が10の8乗ほどあり、また1分子でもよくふえるRNAが出現すると指数増幅するので、確実に組み替え体が現れることになります。
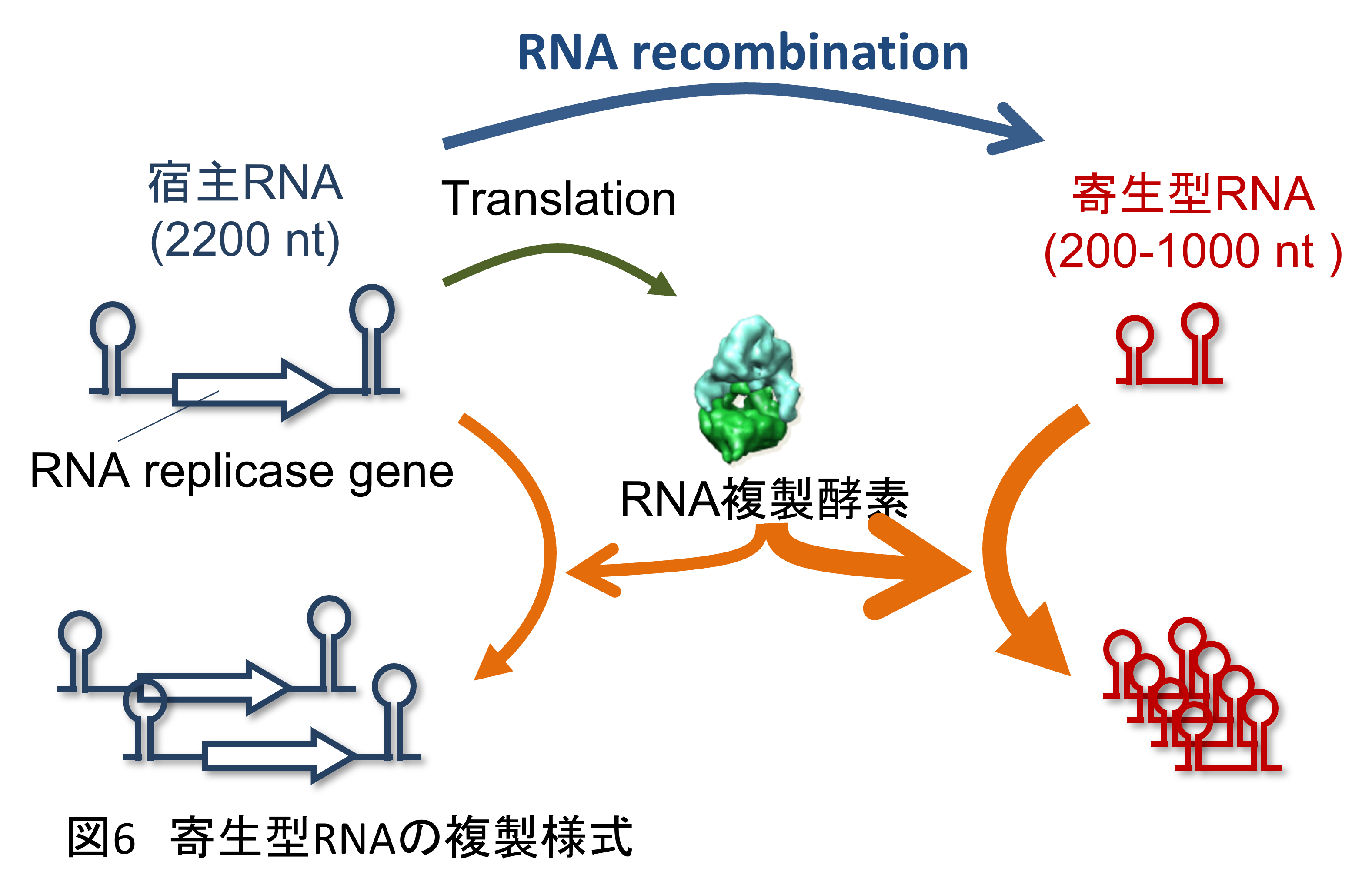
ただ、このような寄生型RNAが発生してしまうと、宿主となるもともとのRNAが増えなくなってしまいます。このために、これまでの実験ではRNAの濃度を低く抑えることで出現しないようにしていたのです。正直なところ迷惑なやつだと思っていたのですが、今やこの迷惑な奴が進化の鍵を握っているかもしれません。そこで、もうRNA濃度を低く抑えることはせず、毎回一定の割合で水滴を次のラウンドに継代するようにしました(4)。そうすると2ラウンド目にはもう寄生型RNAが出現しました(図7)。そしてひとたび出現すると寄生型RNAは短い分、極めて速く増えてしまい、宿主となる元のRNAを圧倒します。そうなると宿主由来の複製酵素が足りなくなり、宿主も寄生体もどちらも増えなくなります。その結果、継代のたびに濃度が低下していくことになりました。しかし、状況が変わるのはある程度寄生型RNA濃度が下がってからです。例えば寄生型RNAの濃度が十分に下がると寄生型RNAの存在しない水滴が現れます。そしてその中では宿主RNAは問題なく増えることができます。そしてそのRNA濃度は今の水滴のサイズでは1 nM程度となります。この実験ではまさにその現象が起こり、寄生型RNA濃度が1 nMを切ったくらいで再び宿主RNAの複製が復活しました。その後を追うように寄生体が再び増え始め、再び複製が止まりました。このように宿主RNAと寄生型RNAは濃度振動を起こしながら複製を続けることがわかりました。
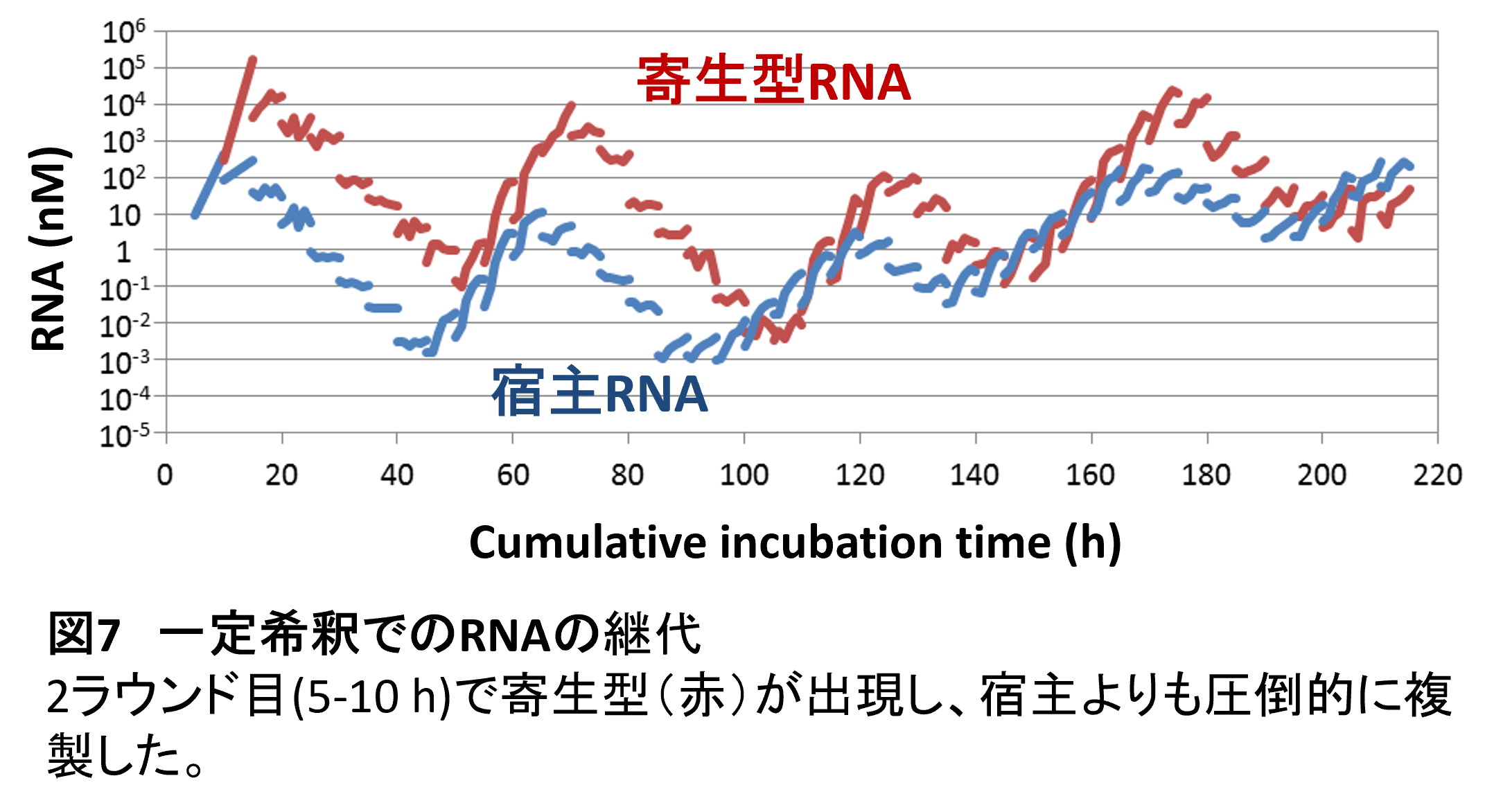
このような濃度ダイナミクス(個体群動態)は、生態系の宿主-寄生体間や捕食者-被食者間で見られるものとよく似ています。しかし、この実験には生物は使われていません。RNAやタンパク質の分子のみです。このような無生物の集合体でも生物と同じ現象を起こすことができるという一つの例です。
これでRNAを寄生体とともに継代することができるようになりましたので、さらに長期にわたって継代してみました(5)。さて、進化は加速するでしょうか? 多様性は生まれるでしょうか?
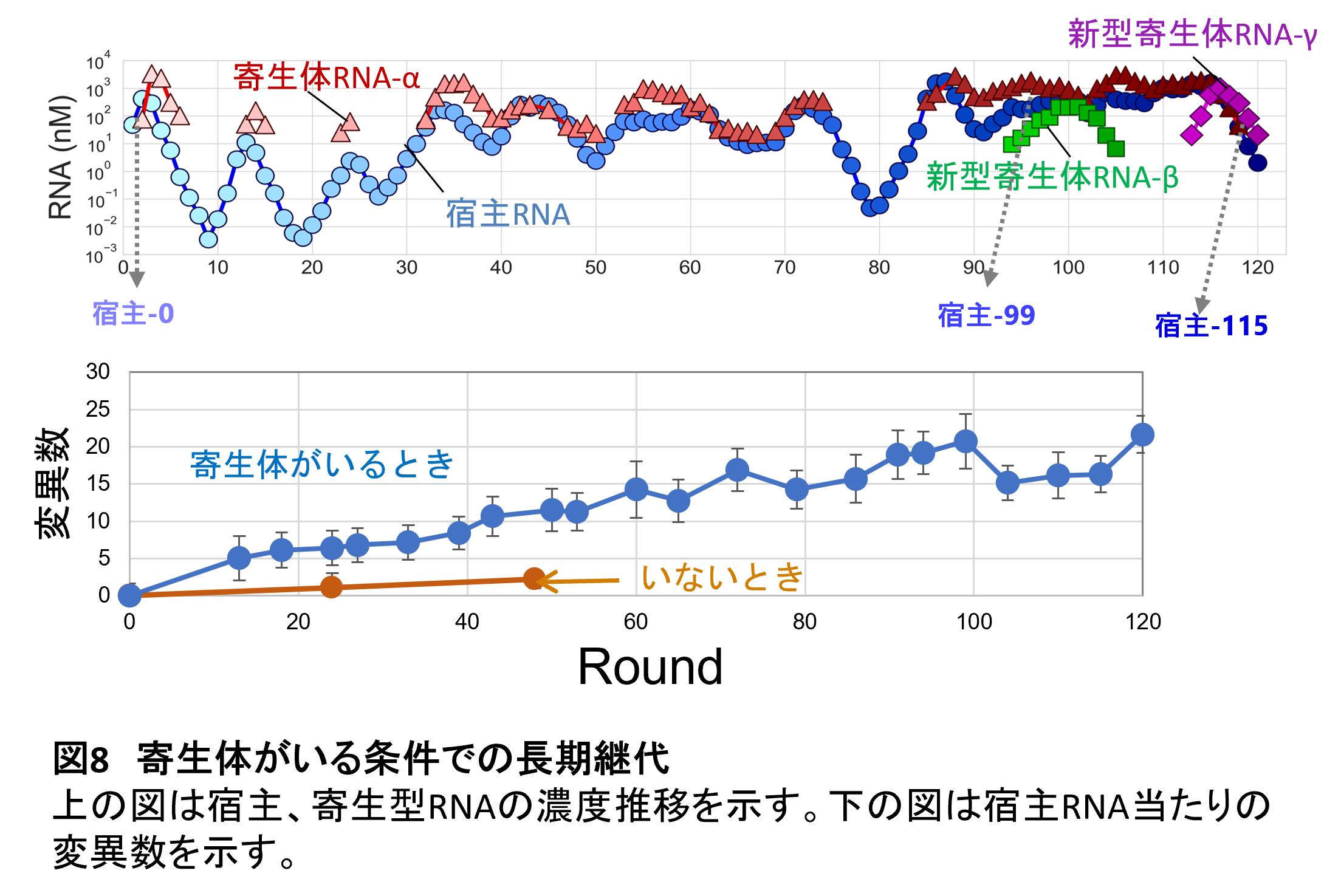
答えはどちらもイエスでした。まず濃度ダイナミクスはめまぐるしく変化しました(図8上)。最初は寄生型RNAに比べて宿主RNA濃度は低く抑えられていたのですが、徐々に宿主RNA濃度が上昇していきました。その後、異なるタイプの寄生型RNAが出現しました。もともとの寄生型RNAは200塩基長の短い物でしたが、後半で出てきたものは約1000塩基と500塩基長という長い(しかし機能のある複製酵素遺伝子は持っていない)ものが出現しました。つまり進化が続いて、寄生型RNAに多様性が生まれてきたことを示しています。
進化が加速したかどうかは宿主RNAに固定された変異の数を比較するとはっきりします(図8下)。この寄生体との共進化実験では、先の寄生体がいない条件で継代した最後のRNAを使っています。したがって、さらに寄生体がいない条件(つまりRNA濃度を低く保った条件)で継代しても、もうほとんど変異が蓄積していきませんが、寄生体と共進化した条件では数倍速く変異が蓄積していきます。つまり進化が加速していることを示しています。
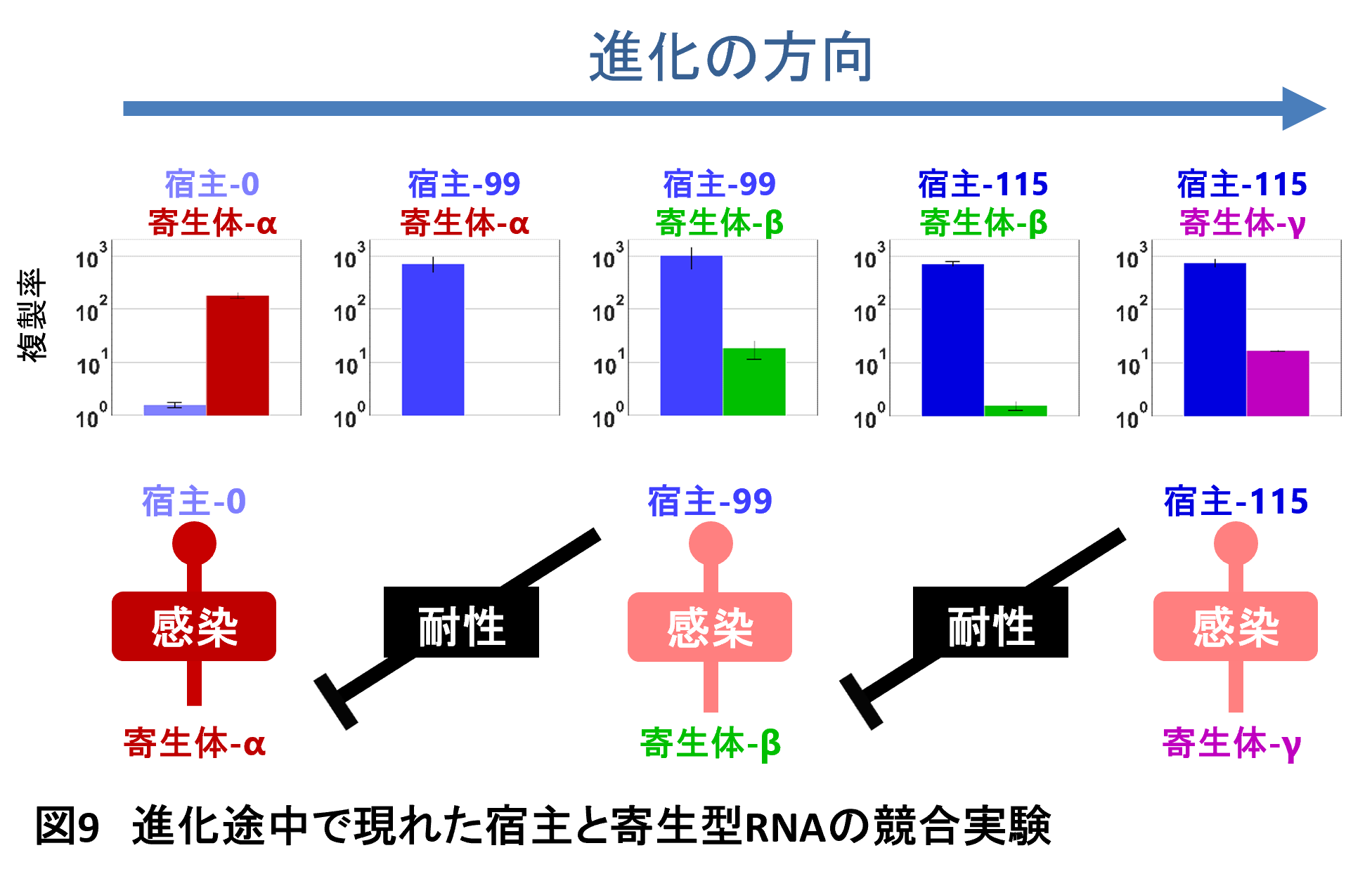
赤の女王仮説によれば、進化の加速は宿主と寄生体との進化的軍拡競争によるはずです。つまり、宿主と寄生体がお互いに適応しあうことによって最適値がどんどん変わっていき、進化が止まらなくなっていると予想されます。これを確かめるために、進化途中に現れた代表的なRNAを宿主と寄生体のそれぞれ3種類選び、競合実験を行いその強さ関係を調べて見ました。この競合実験では宿主と寄生型RNAを1種類ずつ混ぜて、どちらがどのくらい増えやすいかを比べることで強さの関係を測ります。その結果、最初の宿主には最初の寄生体は極めて強かったのが、次に現れた宿主(H99)では力関係が逆転しており、宿主がほぼ完全な寄生体耐性を身に着けていました。しかし、次に現れた寄生体(β)はこの寄生体耐性宿主とも一緒に増えることができるように適応していました。さらに次の最後の宿主はこの寄生体の複製を防ぐ能力を持っていること、さらに最後の寄生体は最後の宿主と一緒に増えられるようになっていることを見出しました(図9)。まさにこの結果は宿主RNAと寄生型RNAがお互いに相手に適応し続ける共進化が起きていることを示しています。
宿主の多様性についてはどうでしょうか? 進化途中のRNA集団について各点で次世代シーケンスにより網羅的にRNA配列を解析して系統樹を描きました(6)(図10)。すると宿主系統は大きく2つの枝へと分岐していることがわかりました。一つの系統(HL1)は変異を蓄積し枝を長く伸ばしています。これは進化が速い系統であることを示しています。一方でもう一つの系統(HL2)の枝の長さは短く、進化があまり進んでいません。しかし、この系統は最後まで維持されていることから(集団内の存在比率は横のヒートマップで示しています)、進化しなくても増え続けられている不思議な系統です。さらにHL2からは上記の新しい寄生型の系統が出現しています(PL2, 3)。これらの寄生型系統はHL1と同程度の枝の長さになっており、HL1と競って進化していることが予想されます。この系統樹の結果は、寄生体との共進化条件では、宿主と寄生体が複数系統へと多様化していることを示しています。
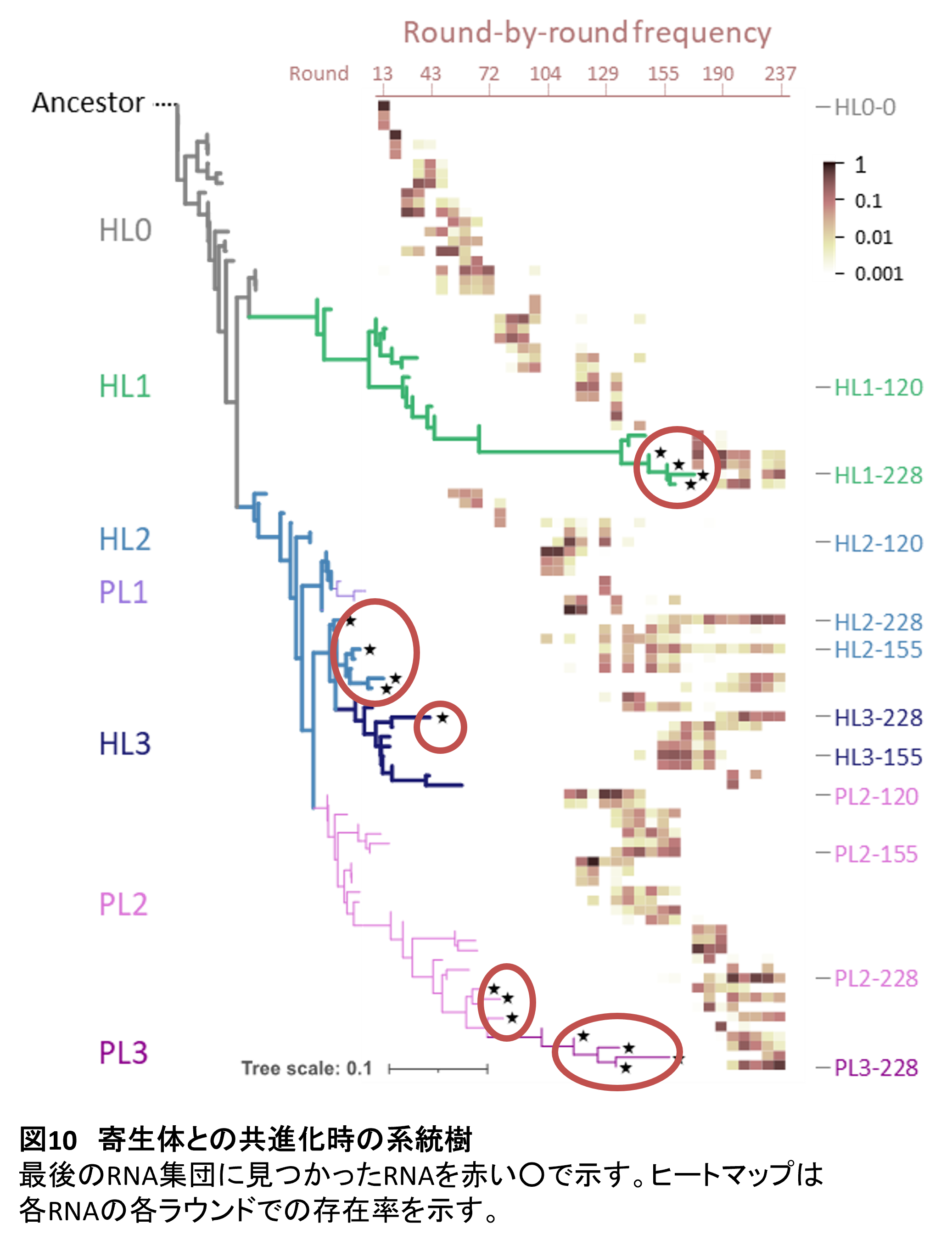
複雑な複製ネットワークの進化
さて、寄生体との共進化により宿主および寄生型RNAが複数系統へと多様化していくことを見出しました。ではこの多様化したRNA達はこの後どうなっていくのでしょうか?
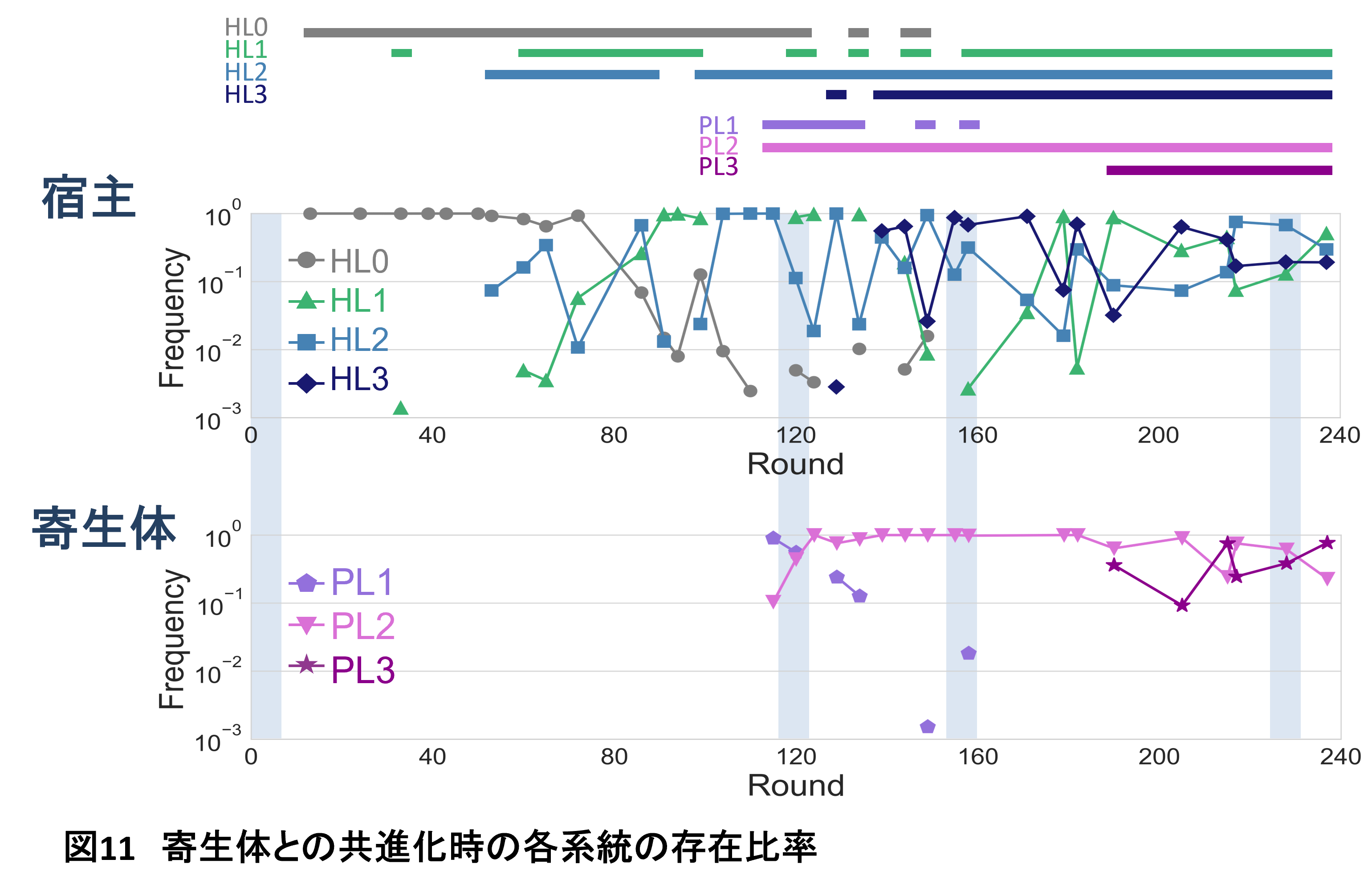
私たちがさらに長期の継代実験を行った結果(図10にはこの結果も一緒に載せています)、もともと多系統は排他的だったのが、だんだん共存するようになってきたことを見出しました(6)(図11)。例えば最初のころの80-160ラウンドではHL1系統の頻度が高くなるとHL2系統の頻度は下がりほとんど検出できなくなります。逆も然りで、つまりこの2つの系統は排他的です。しかし200ラウンドくらいになるとだんだんどちらの系統もある程度の頻度で共存できるようになってきました。つまり、多様化した系統がお互いを排除せずに増えられるようになったことを意味しています。
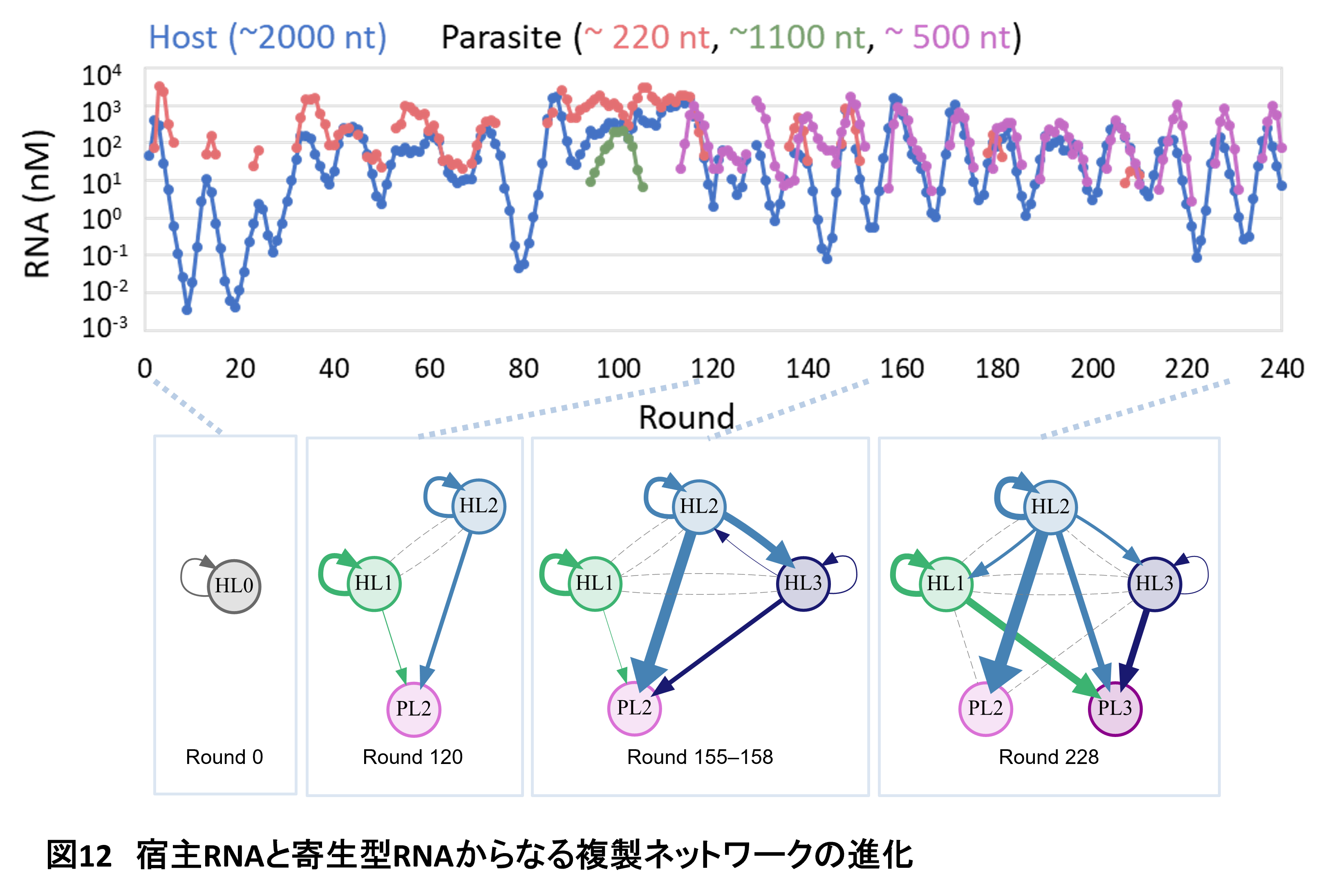
いったいどうやってこんな共存が可能なのかを調べるために、進化途中の各段階で各系統の代表となるようなRNAを単離し、すべての組み合わせでどのRNAがどれくらいどのRNAを増やす能力があるのかを調べました(図12)。RNAの増やしやすさを矢印の太さで示しています。矢印が描かれていないところは増やさないことを示しています。継代を続けるにつれて、RNA系統が増えるとともに、その間の相互作用もだんたん増えていきました。最終的な5種類のRNAについては複雑な複製ネットワークを形成していました。このネットワークの形成するRNAにははっきりした個性がありました。たとえばHL1は宿主RNA系統ですが、自分と特定の寄生型RNAしか増やしません。一方でHL2は自分を含めすべてのRNAを増やすいわば利他的なRNAとなっています。HL3は奇妙な宿主RNA系統で、自分のことをあまり増やさなくなっており、半分寄生体のような性質を持っています。
計算機シミュレーションの結果、このネットワーク構造はHL3を除き安定であることがわかっています。つまり、HL3を除いた4者ネットワークについて、どの1つのRNAを除いても残りの3者ネットワークは維持できません。どれか2つあるいは2つのRNAが複製できずに系から失われてしまいます。つまり、このRNA集団は相互依存的な複製ネットワークを進化させたということを意味しています(HL3については正直なところ謎です。集団中では安定に維持されているのですが…。単離すると状況が違うのかもしれません)。
以上の現在までの長期の共進化実験で分かったことは、「分子を長期進化させると自発的に多様化し複雑な複製ネットワークを形成することができる」ということです。この実験で最初に使ったRNAは1種類の宿主RNAです。私たち実験者が行ったのはただ毎日栄養となる翻訳系を含む水滴を加えて混ぜただけです。何も人為的な選択圧を加えてはいません。それなのに寄生型のRNAが自発的に生まれ、勝手に共進化を始め、多系統へと別れ、ついには複製ネットワークを作って共存し始めました。ここには生きているものは一つもありません。RNAとタンパク質と低分子量化合物のみからできている化学的な反応ですが、寄生体、共進化、多様化、複雑化といった今までは生物でしか起きないとみなされていた生命現象を起こすことができました。実は継代実験はまだ続けており、RNA達はまだ進化し続けています。次に何が起きるのかワクワクしませんか? 私たちのRNA自己複製システムと進化実験により、きっと物質から生命が生まれる条件を明らかにできると期待しています。
もし、興味がある後期教養学部生、大学院生、博士研究員の方がいたら遠慮なく連絡ください。一緒に研究を進めましょう。
参考文献(*は当研究室からの論文)
- *Ichihashi, N.; Usui, K.; Kazuta, Y.; Sunami, T.; Matsuura, T.; Yomo, T. Darwinian Evolution in a Translation-Coupled RNA Replication System within a Cell-like Compartment. Nat. Commun. 2013, 4, 2494.
-
Van Valen, L. A New Evolutionary Low. Evol. Theory 1973, 1, 1-30; Dawkins, R.; Rebs, J. R. K. Arms Races between and within Species; 1979; Vol. 205.
-
Takeuchi, N.; Hogeweg, P. Evolution of Complexity in RNA-like Replicator Systems. Biol. Direct 2008, 3, 1-20; Zaman, L.; Meyer, J. R.; Devangam, S.; Bryson, D. M.; Lenski, R. E.; Ofria, C. Coevolution Drives the Emergence of Complex Traits and Promotes Evolvability. PLoS Biol. 2014, 12, e1002023
-
*Bansho, Y.; Furubayashi, T.; Ichihashi, N.; Yomo, T. Host-Parasite Oscillation Dynamics and Evolution in a Compartmentalized RNA Replication System. Proc. Natl. Acad. Sci. U. S. A. 2016, 113. 4045-4050
-
*Furubayashi, T.; Ueda, K.; Bansho, Y.; Motooka, D.; Nakamura, S.; Mizuuchi, R.; Ichihashi, N. Emergence and Diversification of a Host-Parasite RNA Ecosystem through Darwinian Evolution. Elife 2020, 9
-
*Mizuuchi, R.; Furubayashi, T.; Ichihashi, N. Evolutionary Transition from a Single RNA Replicator to a Multiple Replicator Network. Nat. Commun. 2022, 13, 1460.
世界での立ち位置
この項では上記の私たちの研究が世界の研究でどんな位置を占めているかを(時々聞かれるので)説明したいと思います。
このようなアプローチ(分子の自己複製システムの進化実験)で生命の起源や進化を研究しているのは世界で私たちのグループのみです。独走状態といっても過言ではないと思います。したがって、なにをやっても新規な結果が得られるというかなり恵まれた状況にあります。
他に競合していない理由は、まず第1に、継代実験を実施するのがかなりハードルが高いということがあります。継代実験のためには再構成翻訳系(PURE system)をまあまあ消費します。この翻訳系は市販されていますが、市販品を使うとかなりの出費になりますので私たちは自前で作っています。一度作ってしまえば維持にはそれほど労力はかからないのですが、一から作ろうとするとかなり大変です。また継代実験にも細かいノウハウがあり(特に水滴の調製)、うちのラボメンバーに習えば誰でもできますが、習わずにやろうとすると大抵つまづくような気がします。
第2の理由として、生命の起源や進化という研究領域はもともと解いている問題の大きさに対して研究者が少ないので競争が発生しないという事情もあるのではないかと思います。生命の起源や進化はわからないことだらけですが、あまり実用的ではないと一般に思われているので(後述するようにそうでもないのですが)、研究者人口は多くありません。この状況では、研究者が狭い分野で競争するよりも、お互いに得意分野で研究を進め一緒に理解を深めていく方が効率的です。理想的な科学の進め方のように思います。
また上で「生命の起源や進化の研究はあまり実用的でないと思われている」と書きましたが、個人的には全くそんなことはないと思います。研究者が少なく、手の付けられていない領域には大きなフロンティアがあります。実際に試験管内で分子を進化させた研究は、その後、RNA、DNAリボザイムやアプタマーの人為選択手法として利用されていますし、もともとRNAワールドとタンパク質ワールドをつなぐ分子として探索されたtRNAにアミノ酸をつけるリボザイムは、現在では環状ペプチドの合成に利用されています。生命の起源と進化の分野は物質と生命の境界領域です。そこのは今の生物は利用していないけど今の生物に影響しうる物質が多く眠っていると感じており、今まで誰も予想しなかったブレイクスルーは案外こんなところから出てくるような気がしています。そう感じているのは私だけではなく、生命(とその情報処理機構)の起源の解明に1000万ドルの懸賞金がかけられたりもしています(https://www.herox.com/evolution2.0)。
これからやりたいこと
進化という現象の理解につなげる
今後の一つの方向性として、これまでに観察された進化現象をもたらした原理を理解したいと考えています。
普通の過去の地球で起きた生物進化であれば、なぜその進化が起きたのかは限られたデータ(現存するDNA配列や化石資料)から推測することしかできませんが、私たちの行っている進化実験では進化途中のすべての段階のRNA集団が保存されています。次世代シーケンサーを使えばその集団の配列組成を網羅的に解析することができます。実験条件を変えて途中から進化をやり直すことも自由にできます。つまり進化途中で何が起きていたのか、どの条件がそれを決定したのかをすべて理解することができます。
また私たちの複製系は生物に比べると圧倒的に単純なシステムですので、計算機シミュレーションによってある程度ふるまいを記述することもできます。これにより、どんな条件が整えばどんな進化現象がおきるのかの十分条件を明らかにすることができます(注3)。
現在、例えば以下のようなことを知りたいと思っています。
・どこまで多様化するのか、上限は何で決まっているのか?
・HL1, HL2のような性質の異なる宿主系統が出現するのは必然なのか?
・寄生体との共進化を経験した宿主の方が(寄生体非存在下でも)よく増えるようになっているのはなぜか?寄生体がいると適応度地形の谷を乗り越えられるのか?
・進化実験によって多様性が生まれるときと、生まれない時があるのはなぜか?
・RNA構造揺らぎから進化可能性や方向性を予測できるか?
注3:ときどき聞かれる質問として、私たちの複製系くらい単純であればすべて計算機シミュレーションで再現できてしまうのでは?(実験もしなくていいのでは?)というものがありますが、そんなことはありません。どうしても計算機でシミュレーションできない要素は、変異の影響です。進化の起点となるのは変異です。進化をシミュレートするためにはRNAに変異が入ったときにRNAの性質がどう変わるかを予測できないといけませんが、その予測ができません(それは変異が入ったときにRNAやタンパク質の構造がどう変わって、その構造変化が生化学的な性質をどう変えるのかが未だ予想できないことに起因します)。したがって、現状では進化実験なしに信頼に足る進化シミュレーションはできません。計算機シミュレーションが役に立つのは、出現しうるRNAやタンパク質の性質が分かっている場合に限られます。
継代の自動化
今後、さらなる継代実験を行いたいと思っていますが、問題となるのは継代にかかる時間です。今まで起きなかった現象を見たいと思うと、経験上、これまでの継代回数の倍の数だけ継代する必要がありますが、次第に労力が許容できないレベルになっていきます。そこでそろそろ自動化したいと考えています。実は以前に自動継代システムを作っていたのですが、規模が大きすぎて並列で複数継代できないという問題と、反応時間や継代量の調整ができないなどの問題がありました。そこでもっと簡単で反応時間や植え継ぎ率を制御できるような自動継代システムを作りたいと思っています。こうした工作ができるような人材を切に求めています。
上記のテーマ以外でも、新しいテーマは日々生まれていますし、面白く生命の理解を高めるテーマは大歓迎です。一緒にやりたい! という学生、PDの方は一度研究室に来てみてください! 学振PDも大歓迎です。
詳しくはこちら
この研究で身につくこと
もし、大学院、あるいは博士研究員として上記のような研究をした場合、何よりまず未知の現象から人間に理解できる要素を抽出する能力が身に付きます。進化実験では、基本的に予想したことは起きません。これは私たちの知識が全然足りていないためだと思います。何か複製能力の高いRNAは必ず取れてきますが、なぜそのRNAが複製しやすいのかがよくわからないことがほとんどです。それを多方面のデータから仮設を立てて、たくさんの実験を行い、仮説を絞り込んでいく作業が必要になります。その中でぐちゃぐちゃした自然現象を抽象化して理解するための腕が磨かれていくはずです。これは自然科学を行う上で最も重要な能力の一つで、卒業後に何の研究を行うにしろ役に立つ能力が得られるはずです。また卒業後に研究をしなかったとしても、この複雑な社会で価値のあるものを作っていくためには、同じ能力はきっと役に立つはずです。
上記のような抽象的な能力が大学院で身につく大事な能力だと思いますが、それ以外にももう少し具体的なスキルとしては、基本的なDNAやRNAの取り扱い技術、大量の配列データを解析するバイオインフォマティクス技術、計算機シミュレーション技術なども身に着けることができます。